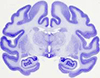
These include: the medial preoptic nucleus; median and lateral preoptic area; medial division of the bed nucleus of stria terminalis; paraventricular nucleus; central nucleus of the amygdala; dorsal hypothalamic area/dorsomedial hypothalamus; lateral hypothalamic area; lateral, ventrolateral and dorsomedial divisions of the periaqueductal grey; dorsal raphe nuclei; parabrachial nuclei; Kölliker-Fuse nucleus; intertrigeminal region; rostral ventrolateral medulla; lateral parafacial region; and the ventral respiratory group. 

Plantar CVC responses were evoked most strongly from the dorsal hypothalamic area and most dorsal part of the dorsomedial nucleus, whereas peak phrenic rate responses were evoked from more caudal sites; their relative magnitudes varied systematically with rostrocaudal position.
The HGA includes parts of the paraventricular nucleus and the dorsal hypothalamic area.
More recently, we demonstrated that (1) the tachycardia resulting either from chemical stimulation of the DMH or from experimental stress is markedly attenuated by microinjection of the GABAA receptor agonist muscimol, a neuronal inhibitor, into the medullary raphe pallidus (RP); and (2) the specific subregion of the DMH mediating stimulation-induced tachycardia corresponds to the dorsal hypothalamic area (DHA), a site where neurons projecting to the RP are densely concentrated.
Among injections of cholera toxin b-subunit (CTb), a retrograde tracer, into hypothalamic regions at the rostrocaudal level of the DMH, injections into the DMH, lateral hypothalamic area (LH) and dorsal hypothalamic area (DH) resulted in EP3 receptor immunolabelling in substantial populations of CTb-labeled neurons in the POA.
RESULTS: We found that Narp colocalizes with hypocretin in the lateral hypothalamic area (LHA), the dorsomedial hypothalamus (DMH), the dorsal hypothalamic area (DHA), and the posterior hypothalamic area (PHA) of the normal human.
Microdialysis to the dorsal hypothalamic area was conducted in the same animal before and after cholestasis. Delta- and kappa-stimulated binding was significantly decreased in cholestasic animals compared to controls in the dorsal hypothalamic area.
On the other hand, when cholera toxin-b was injected into the caudal periaqueductal gray matter, many double-labeled cells were seen in a cell group extending from the dorsomedial nucleus through the dorsal hypothalamic area in cold-exposed rats but few were seen in warm-exposed rats. These results suggest that the rostral periaqueductal gray matter receives input from the median preoptic nucleus neurons activated by warm exposure, and the caudal periaqueductal gray matter receives input from neurons in the dorsomedial nucleus/dorsal hypothalamic area region activated by cold exposure.
In the hypothalamus, irNPB cells were present in the medial preoptic area and nucleus, ventromedial preoptic nucleus, retrochiasmatic nucleus, paraventricular hypothalamic nucleus, supraoptic nucleus, accessory neurosecretory nuclei, periventricular hypothalamic nucleus, dorsomedial hypothalamic nucleus, supraoptic retrochiasmatic nucleus, lateral hypothalamic area, posterior hypothalamic area, dorsal hypothalamic area, and zona incerta.
Saline stimulated FOSir in the paraventricular thalamic nucleus (PVA) and the dorsal hypothalamic area (DA) in MKO mice, but not in wild-type (WT), suggesting that MKO homozygotes may differ responsively from WT.
Distinct clusters of CRF-BP-ir neurones were identified in the anterior and posterior parvocellular and dorsal cap subdivisions of the paraventricular nucleus (PVN), as well as in the dorsal hypothalamic area, dorsomedial hypothalamic nucleus (DMN), ventral premammillary nucleus and zona incerta.
In the rat, Y1-R mRNA expression was also seen in the subfornical organ, anterior hypothalamic area, dorsal hypothalamic area, and in the lateral hypothalamic area.
No significant variation in the MTNR1A mRNA expression was observed in a more dorsal hypothalamic area.
Neurons projecting to the RP from the DMH are said to be concentrated in a specific subregion, the dorsal hypothalamic area (DA).
Compared to control cats, c-Fos immunoreactive cells were significantly increased (P<0.05) in the arcuate nucleus (ARC), dorsal hypothalamic area (HDA), dorsomedial nucleus, paraventricular hypothalamic nucleus (PVN) and periventricular nucleus in the BK-treated animals.
Microinjections outside the DMH within the dorsal hypothalamic area adjacent to the mamillothalamic tracts or within the ventromedial hypothalamus were less effective for attenuating the PGE2-evoked thermogenic, metabolic, and cardiovascular responses.
In addition, some scattered bNOS labeled somata were noted in the dorsal hypothalamic area.
Long-term undernutrition enhanced the number of galanin neurones located in the infundibular nucleus and the dorsal hypothalamic area (DHA), refeeding resulted in an increase of neurones in the DHA and preoptic area, but short-term starvation had no effect on any galanin subpopulations.
These effects were confined to a specific region that included medial portions of the dorsal hypothalamic area and dorsal ependymal, subependymal, and neuronal components of the periventricular nucleus.
There were no qualitative differences in the distribution of proliferation zones between the sex phases studied with the exception of the ventricular region of the dorsal hypothalamic area.
There were novel groups of neurotransmitter- or synthetic enzyme-immunoreactive neurons such as tyrosine hydroxylase-immunoreactive cells in the bed nucleus of the stria terminalis, dopamine-beta-hydroxylase-immunoreactive cells in the motor trigeminal, hypoglossal, and paraabducens nuclei, and arginine vasopressin- and oxytocin-immunoreactive cells within the nucleus of the diagonal band, dorsal hypothalamic area, and arcuate nucleus compared with Syrian hamsters.
with CL316243 compared with control rats in the paraventricular hypothalamus, lateral hypothalamic area, ventromedial hypothalamic nucleus and dorsal hypothalamic area.
Other hypothalamic subdivisions displaying numerous radiolabeled cells were the dorsomedial, ventromedial, infundibular and tuberomammillary nuclei, and the dorsal hypothalamic area.
Moderate levels of GALR2 mRNA are observed in the olfactory bulb, olfactory tubercle, piriform and retrospinal cortices, hypothalamus (namely the preoptic area, arcuate nucleus, and dorsal hypothalamic area), substantia nigra pars compacta, and sensory trigeminal nucleus.
Many neurons in the posterior (A11) and caudal dorsal hypothalamic areas (A13) as well as in the arcuate (A12) and periventricular (A14) zone were immunoreactive for the two enzymes, suggesting that they were dopaminergic. Numerous tyrosine hydroxylase-immunoreactive neurons, which were not immunoreactive for aromatic L-amino acid decarboxylase, could be seen in the paraventricular, supraoptic and accessory nuclei (A15) as well as in the rostral dorsal hypothalamic area.
NADPH-d positive and/or ASS-immunoreactive neurons were localized in the following areas: the anterior hypothalamic area, the anterior hypothalamic nucleus, the supraoptic nucleus, the suprachiasmatic nucleus, the periventricular complex, the paraventricular nucleus, the parvocellular nucleus, the lateral hypothalamic area, the dorsomedial hypothalamic nucleus, the dorsal hypothalamic area, the posterior hypothalamic area, and the supramammillary nucleus.
In agreement with previous retrograde studies, after all injection sites, a substantial to large number of labeled neurons were observed in the dorsal hypothalamic area and dorsolateral and ventrolateral parts of the periaqueductal gray, and a small to moderate number were found in the lateral preoptic area, bed nucleus of the stria terminalis, paraventricular hypothalamic nucleus, central nucleus of the amygdala, lateral hypothalamic area, parafascicular area, parabrachial nuclei, subcoeruleus area and parvocellular reticular nucleus.
Inhibition of C-responses could also be produced by stimulation of the dorsal hypothalamic area (DA).
The present results confirm previously reported afferent connections of the infralimbic area and further reveal additional inputs from the anterior amygdaloid area, anterior cortical amygdaloid nucleus, posterolateral cortical amygdaloid nucleus, ventral reunions, lateral preoptic area, dorsal hypothalamic area, posterior hypothalamic nucleus, supramammillary nucleus and medial mammillary nucleus.
In the diencephalon, both markers were found in the lateral hypothalamus, dorsal hypothalamic area, hypothalamic periventricular nucleus, paraventricular nucleus and mamillary area.
Other hypothalamic nuclei that contain endotoxin-induced Fos nuclear immunostaining include the parvocellular neurons of the paraventricular nu., the dorsomedial and arcuate nuclei, the lateral hypothalamus, the dorsal hypothalamic area (zona incerta), and the median nucleus of the preoptic area.
The amplitude of the depressor responses was significantly correlated with baseline MBP on stimulation of the dorsomedial hypothalamic nucleus (DMH) or the dorsal hypothalamic area.
In the hypothalamus, in addition to the perifornical region, retrogradely labeled neurons were found in all cases in the tuberomammillary nucleus, premammillary nucleus, dorsal hypothalamic area, ventromedial hypothalamic nucleus, and the paraventricular nucleus.
Moderate densities were found in the vertical limb of the diagonal band of Broca, the bed nucleus of the stria terminalis, the ventral part of the medial preoptic area in neonate/infant, the medial and lateral mammillary nuclei in neonate/infant, the supramammillary nucleus in adult, the dorsal hypothalamic area, and the ventromedial nucleus.
Although numerous TRH-immunoreactive (TRH-i) neurons are present in other hypothalamic areas of the brain, only a few of them in the dorsal hypothalamic area behind the PVN and the periventricular preoptic nucleus could be retrogradely labeled.
This hypothalamic "grooming area" consists of parts of the hypothalamic paraventricular nucleus and of the dorsal hypothalamic area.
The histaminergic projection appears to be separate from a previously reported, apparently non-histaminergic projection from neurons in the dorsal hypothalamic area to discrete regions of intermediate and deep colliculus.
Numerous long HA-ir axons coursed dorsomedially from the ventrolateral posterior hypothalamus to the dorsal hypothalamic area.
There was little labelling in the dorsal hypothalamic area but moderate labelling in the posterior hypothalamic area.
The results of the study showed that neurokinin B peptide-2-like immunoreactive neurons were located in the nucleus arcuate, median eminence, ventral and external bed nuclei of the stria terminalis, dorsal hypothalamic area, and medial habenula. Neurokinin B peptide-2 neurons that give rise to the long descending projections from the hypothalamus to thoracolumbar spinal cord were found only in the dorsal hypothalamic area. Approximately 36% of the neurokinin B peptide-2 neurons in the dorsal hypothalamic area projected to the spinal cord, whereas about 28% of the spinal projecting neurons in the dorsal hypothalamic area contained neurokinin B peptide-2-like immunoreactivity. Most of the spinal projecting neurokinin B peptide-2 neurons in the dorsal hypothalamic area had a cell size of 15 x 25 microns.
The density of labeled terminals was especially high in the agranular insular cortex, olfactory tubercle, medial division of the mediodorsal nucleus of the thalamus, dorsal hypothalamic area and the lateral division of the central amygdaloid nucleus.
Cocaine also decreased NT binding in non-mesocorticolimbic areas, including the dorsal hypothalamic area and the zona incerta, but binding returned toward control levels 10 days after withdrawal from cocaine.
Electrical stimulation of the hypothalamic paraventricular nucleus (PVH) and of the adjacent dorsal hypothalamic area (DHA) evokes grooming behaviour.
In the hypothalamus, these areas included the lateral hypothalamus (including the medial forebrain bundle), the posterior hypothalamic area, the dorsal hypothalamic area, the dorsomedial nucleus, the paraventricular nucleus, the periventricular area, the suprachiasmatic nucleus, and the lateral and medial preoptic areas.
The largest number of gastrin releasing peptide-immunoreactive axons leaving the suprachiasmatic nucleus could be followed in a caudodorsal direction to the subparaventricular zone and the dorsal hypothalamic area.
The dorsal hypothalamic area, the paraventricular and medial mammillary nuclei displayed low but measurable densities.
Furthermore, labeled neurons were found in the contralateral medial pretectal area in guinea-pigs, in the ipsilateral tegmental mesencephalic reticular formation in monkeys, in the laterodorsal tegmental nucleus in rabbits, and in the dorsal hypothalamic area near the nucleus of the anterior commissure in monkeys.
Injections in the dorsal hypothalamic area produced a similar small response.
Other areas that contained double-labeled cells were: the medial preoptic area, bed nucleus of the stria terminalis, periventricular hypothalamic nucleus, lateral hypothalamic area, dorsal hypothalamic area and subincertal nucleus.
Following HRP injection into the LS, double-labeled cells which contained a homogeneous blue reaction product of ENKI and a black or brown granular reaction product of retrogradely transported HRP were identified in 4 discrete brain regions: perifornical hypothalamic area at the level of the paraventricular nucleus (PeF); posterior part of the anterior hypothalamic nucleus (AHP); bed nucleus of the stria terminalis, medial division, posterolateral part (BSTMPL); and dorsal hypothalamic area (DA).
Furthermore, when 5-hydroxytryptophan was injected with inhibitor of monoamine oxidase, a large number of small neurons immunoreactive to serotonin was identified in many discrete regions: the anterior and lateral hypothalamic areas, preoptic area, suprachiasmatic nucleus, dorsal hypothalamic area, dorsomedial nucleus, posterior hypothalamic area and nucleus of the fields of Forel.
After the iontophoretic application of PHA-L to the dorsal hypothalamic area, labeled fibers that finally ended in the nucleus raphe pallidus were observed descending through the most medial part of the ventral tegmental area and the nucleus reticularis tegmenti pointis to reach the medial aspect of the pyramid. These results strongly indicate that neurons in the dorsal hypothalamic area have a direct connection with neurons in the nucleus raphe pallidus and the ventral part of the PGCL.
In the second set of experiments, ipsilateral double-labelled cells which contained both retrogradely transported horseradish peroxidase (HRP) and CRFI were observed in the zona incerta, subincertal nucleus, lateral hypothalamic area, perifornical hypothalamic area, and in the dorsal hypothalamic area after unilateral HRP injections into the cerebral cortex.
Finally, in the dorsal hypothalamic area as many as 55-75% of the neurons with spinal projections are dopaminergic, on the basis of their staining with an antiserum against tyrosine hydroxylase.
They were identified in the posterior hypothalamic area, rostral arcuate nucleus of the hypothalamus, dorsal hypothalamic area, and periventricular complex of the hypothalamus, which contain tyrosine hydroxylase (TH)-IR cells and are known as A11 to A14 dopaminergic cell groups. AADC-IR perikarya were also found in the other hypothalamic areas where few or no TH-IR cells have been reported: the supramamillary nucleus, tuberomamillary nucleus, pre- and anterior mamillary nuclei, caudal arcuate nucleus, dorsal hypothalamic area immediately ventral to the mamillothalamic tract, anterior hypothalamic area, area of the tuber cinereum, retrochiasmatic area, preoptic area, suprachiasmatic and dorsal chiasmatic nuclei.
The dorsal hypothalamic area appears to be the main target of the efferents to the hypothalamus.
The present study was undertaken using retrograde labeling techniques to clarify whether the neurons in the dorsal hypothalamic area (DHA) that project to the spinal cord are the same as those that project to the nucleus raphé pallidus (NRP).
dorsomedialis-projection to the dorsal hypothalamic area.
The main finding of this study is the disclosure of the fact that the entopeduncular nucleus as well as other cell groups as dissimilar as the lateral preoptic area, the central, medial and anterior cortical amygdaloid complex and neurons of the dorsal hypothalamic area appear to be embryologically related, as they are all derived from a common longitudinal slab of the matrix of the forebrain..
Although the dorsal hypothalamic area contains the largest number of labeled hypothalamic neurons, labeled cells are also found within the periventricular, paraventricular, dorsomedial, ventromedial, posterior, lateral, and anterior hypothalamic nuclei. Following the placement of injections of tritiated anterograde tracers within the dorsal hypothalamic area, transported label is organized in two bands of clusters over the SGI.
Projections arise also in the lateral rostral and caudal parts of the reticular thalamic nucleus, the zona incerta, the lateral and dorsal hypothalamic area, the claustrum, and the entopeduncular nucleus.
The ventral periventricular projection follows the ventral component of the third ventricle into the hypothalamus, passing primarily through the dorsal hypothalamic area and labeling the rostral hypothalamus and preoptic area.
In addition a small cluster of parvocellular vasopressin neurons was present adjacent to the top of the third ventricle in the posterior dorsal hypothalamic area and a number of such neurons were found in the ventral locus coeruleus and sub coeruleus.
In the diencephalon these regions included the supraoptic nucleus and the dorsal hypothalamic area, extending into the caudal portion of the paraventricular nucleus.
In addition, retrogradely filled cells were present in ventromedial, dorsomedial, and dorsal hypothalamic nuclei and in the dorsal hypothalamic area.
The lateral group of cells emanating from the zona incerta occupies the lateral part of the dorsomedial and anterior hypothalamic nuclei and the dorsal hypothalamic area.
Microinjection of bombesin into a variety of brain regions demonstrates that this peptide's site of action is within the dorsal hypothalamic area. Microinjection of the somatostatin analog, desAA1,2,4,5,12,13 [ D-Trp8]-somatostatin (ODT8-SS) into the dorsal hypothalamic area but not into other brain areas prevents bombesin-induced elevation of plasma glucose and epinephrine.
Projections from the A1 region ascend through the lateral hypothalamus to the bed nucleus of the stria terminalis, the medial preoptic area, and several hypothalamic nuclei: the dorsomedial nucleus, the dorsal hypothalamic area, the paraventricular and supraoptic nuclei, and the median eminence.
Cells of origin of this novel projection were identified with the retrograde horseradish peroxidase (HRP) method in the dorsal hypothalamic area at the most rostral level of the dorsomedial hypothalamic nucleus..
The dorsal hypothalamic area and supramammillary nucleus show restricted projections to the central and basolateral nuclei, respectively.
After injections of an aqueous solution of horseradish peroxidase (HRP) restricted to the superior colliculus of the cat, labeled neurons were found ipsilaterally in the dorsal hypothalamic area, lateral hypothalamus, zone incerta, reticular thalamic nucleus and ventral lateral geniculate.
Cells labeled with the peroxidase reaction product were located in gyrus propreus, gyrus genualis, nucleus accumbens, bed nucleus of the stria terminalis, bed nucleus of the anterior commissure, nucleus of the diagonal band, substantia innominata, anterior amygdaloid area, ventromedial hypothalamic area, paraventricular nucleus, perifornical hypothalamic area, lateral hypothalamic area, dorsal hypothalamic area, field of Forel, midbrain reticular formation, superior colliculus, ventral central grey, lateral central gray, locus coeruleus, parabrachial nuclei, nucleus of the lateral lemniscus, oral pontine reticular nucleus, and the dorsal raphe.
Cobalt filled fibres were found in the preoptic area, dorsal hypothalamic area, and in the periventricular region close to the wall of the third ventricle..
The ACTH inhibitory area appears to be an extension of portions of the central tegmental tract and to extend medially to the posterior hypothalamic area and the dorsal hypothalamic area and ventrally toward the basal hypothalamus.
Other moderately to intensely stained AChE neurons were found in the lateral and perifornical areas and most particularly in the dorsal hypothalamic area.
One division of fibers continues forward beneath the ventral thalamic group and distributes to the zpna incerta and dorsal hypothalamic area.
The heaviest accumulations of catecholamine-containing varicosities were seen within the: anterior periventricular nucleus; dorsal hypothalamic area; bed nucleus of the inferior thalamic peduncle; doral component of the paraventricular nucleus; dorsomedial nucleus; infundibular nucleus; bed nucleus of the stria terminalis-medial division; and supraoptic nucelus.
-
[ View All ]